
Kambrikausi
| Kambrikauden jako | ||||
|---|---|---|---|---|
| kausi | epookki | vaihe | ikä (mvs) | |
| Ordoviki | varhais | Tremadoc | nuorempi | |
| Kambri | Furong | 10. vaihe | 488,3–492,0 | |
| Jiangshan | 492,0-496,0 | |||
| Paibi | 496,0–499,0 | |||
| 3. sarja | Guzhang | 499,0–503,0 | ||
| Drum | 503,0-506,5 | |||
| 5. vaihe | 506,5–510,0 | |||
| 2. sarja | 4. vaihe | 510,0–515,0 | ||
| 3. vaihe | 515,0-521,0 | |||
| Terreneuve | 2. vaihe | 521,0-528,0 | ||
| Fortune | 528,0–542,0 | |||
| Ediacara | vanhempi | |||
| Kambrikauden jako ICS:n mukaan.[1] Kansainvälinen kerrostumajärjestö ei ole antanut virallista nimeä monellekaan kambrikauden kerrostumalle. Nämä ovat järjestyksellisiä vaiheita väliaikaisesti. | ||||

Kambrikausi oli geologinen ajanjakso 542–490 miljoonaa vuotta sitten. Se oli monisoluisten kuorellisten merieläinten nopeaa kehittymisaikaa. Kaudella ei ollut vielä maaeläimiä. Uusia eläinryhmiä ja lajeja syntyi sekä kuoli nopeasti. Kambrikauden eliöt olivat enimmäkseen ajan lämpimissä ja matalissa merissä eläneitä selkärangattomia, kuten nilviäisiä sekä molukkirapua muistuttaneita trilobiitteja. Eläimille kehittyi suojakuoria pedoilta puolustautumiseksi. Aikakauden nopeaa lajikehitystä kutsutaan kambrikauden lajiräjähdykseksi. Kambrikauden melko suurikokoisilla kovakuorisilla eliöillä oli ollut pienempiä, pehmeäkuorisempia edeltäjiä. Kauden alussa eli nykyisin tuntemattomia merieläimiä.
Nimen tausta
[muokkaa | muokkaa wikitekstiä]Kambrikausi on saanut nimensä Walesin latinankielisestä nimestä Cambria. Englantilainen geologi Adam Sedgwick nimesi Etelä-Walesissa ja Lounais-Englannissa tutkimiensa liuskekivikerrostumien kerroksen vuonna 1835 kambriseksi systeemiksi (engl. Cambrian System). Siihen kuuluvat kambrikaudella syntyneet tai kerrostuneet kivet.[2][3] Virallisesti nimi kambrikausi hyväksyttiin käyttöön vuonna 1960.[3]
Maantiede ja ilmasto
[muokkaa | muokkaa wikitekstiä]
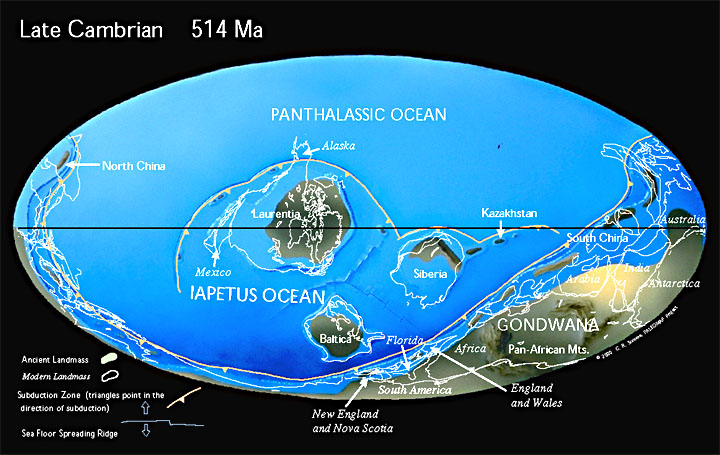
Kambrikauden mantereet erosivat suuresti nykyajan mantereista. Geologien arvioiden mukaan 30. eteläisellä leveysasteella sijaitsi jättiläismanner Gondwana.[4] Mannerta ympäröi mittavan kokoinen, pohjoisnavalle ulottunut valtameri Panthalassa. Osana Gondwanaa olivat nykyisistä alueista Afrikka, Intia, Arabia, Etelä-Amerikka, Etelämanner ja Australia.[5] Nykyinen Aasia oli pirstaleinen. Gondwanan itäpuolella sijaitsi lukuisia pikkumantereita kuten Laurentia ja Baltica sekä nykyisten Siperian, Kazakstanin ja Kiinan alueet. Laurentia toimi nykyisten Grönlannin ja Pohjois-Amerikan alkulähteenä. Muun muassa nykyinen Suomi oli osana Balticaa. Se sijaitsi noin 50. eteläisellä leveysasteella.[6] Gondwana vaikuttaa ajautuneen osittain navan alueelle myöhäiskambrikauden aikana.[7]
Kambrikausi alkoi jääkauden päättyessä. Kauden ilmasto oli leuto sekä nykyistä lämpimämpi ja kosteampi. Jääkauden arvellaan tuhonneen aikaisempaa lajistoa ja antaneen täten tilaa uusien eliöiden nopealle kehitykselle. Lämpimät sekä matalat meret olivat yleisiä kambrikaudella. Merenpinta on ollut luultavasti korkealla ja mannerliikunnot ovat olleet nopeita.
Hieman ennen kambrikauden alkamista, noin 580 miljoonaa vuotta sitten syntynyt jättiläismanner Pannotia oli alkanut jo hajota kauden alkupuolella. Ilmakehässä oli kambrikauden alussa jo huomattava määrä happea, josta oli syntynyt maankamaraa suojaava, mahdollisesti evoluutiota kiihdyttänyt otsonikerros. Kauden aikana tapahtui viisi pientä, alle 30 prosenttia lajistosta pyyhkäissyttä joukkotuhoa. Niistä viimeinen tuhosi 50 prosenttia helmiveneistä ja trilobiiteista sekä päätti kambrikauden. Joukkotuhoa seurasi jääkausi.
Ilmakehän hiilidioksidipitoisuus kohosi kambrikauden alkupuolella 4 500 ppm:stä 7 000 ppm:ään, mutta palautui ennalleen kauden loppuun mennessä, ollen silloinkin huomattavasti nykyistä korkeampi. Toisen tutkimuksen mukaan hiilidioksidipitoisuus olisi ollut läpi kambrikauden suunnilleen 15 kertaa nykyistä suurempi. Planeetan keskilämpötila oli kambrikaudella 5,0–5,5 celsiusastetta nykyistä korkeampi ja yhteensä noin 22 astetta. Maa oli kambrikaudella lämmin, koska päiväntasaajan seudulla kiersi lämmittävä merivirta.
Kasvillisuus ja eläimistö
[muokkaa | muokkaa wikitekstiä]Maakasveja ei esiintynyt vielä kambrikaudella. Merissä eli varhaisia leväryhmiä.
Elämä sijaitsi kambrikaudella enimmäkseen vedessä. Maaelämää ei ollut kehittynyt vielä ainakaan mainittavasti. Äyriäisten sukuiset trilobiitit olivat yleisiä. Simpukoiden ja kotiloiden sukulaisia oli myös runsaasti. Monet kambrikauden eliöt olivat kuorellisia mustekaloja. Trilobiitit, esimerkiksi laji Bathyriscus formosis muistuttivat hieman nykyisiä siiroja. Niiden koko vaihteli alle millimetristä 72 senttimetriin.
Kuorelliset merieläimet yleistyivät nopeasti kambrikaudella. Ajan eläimistöön lukeutui jaokkeisia matoja, simpukoita, kotiloita, simpukoita muistuttavia lonkerojalkaisia, piikkinahkaisia, sienieläimiä, sienieläinten sukupuuttoon kuolleita sukulaisia archaeocyatideja, alkeellisia graptoliitteja sekä merililjoja. Tommotia oli merihiiren kaltainen merenpohjalla elävä nilviäinen. Kauden kookkain otus oli 60 senttimetrin mittainen, jonkin verran katkarapua sekä rauskua muistuttava Anomalocaris. Alkeellinen, ankeriasmainen Pikaia oli luultavasti selkärankaisten kantamuoto. Odontogriphuksen kaltaiset eläimet nauttivat ravinnokseen runsaasti mikrobimatoja. Odontogriphus oli litteän muotoinen matomainen eläin, jolla oli ympyrämäinen suu päänsä alapinnalla.[8]
Edeltänyt lajikehitys ja Tommot-Atdaba-vaiheet
[muokkaa | muokkaa wikitekstiä]Pienikuorisia eliöitä tunnetaan jo kambrikautta edeltäneen ediacarakauden lopulta noin 565–543 miljoonan vuoden takaa. Niihin lukeutui muun muassa vendotaenideja, sabellitideja sekä muita putkilomaisia otuksia. Ensimmäiset alkeelliset nilviäiset ilmestyivät kambrikauden alussa. Kovakuorisia lajeja kehittyi noin 5–15 miljoonaa vuotta kestäneessä vaiheessa 543–530 miljoonaa vuotta sitten. Lähes kaikki ediacarakauden eliöt kuolivat kyseisenä aikana sukupuuttoon ensimmäisten petojen ilmestyttyä. Jäljelle jäi vain kuorellisia otuksia, jotka pystyivät suojautumaan saalistajilta. Ensimmäinen tiedetty petoeläin oli nuolimatoja muistuttava Protohertzina.
Melkein kaikki nykyisin tunnettavat kovakuoristen pääjaksot ilmestyivät Tommot- ja Atdaba-vaiheiden aikana 530–525 miljoonaa vuotta sitten. Seuranneiden 10–20 miljoonan vuoden aikana ilmestyi lukuisia kuoriaisia, petoja, kuppimaisia archaeocyanteja, sienieläimiä, kalkkileviä ja korallieläinten kaltaisia eliöitä. Noin 530–527 miljoonaa vuotta sitten Tommot-vaiheella kehittyivät lonkerojalkaiset sekä monia pehmeäruumismaisia eliöitä. Trilobiitit sekä alkeelliset piikkinahkaiset ilmestyivät noin 527–525 miljoonaa vuotta sitten Atdaba-vaiheen aikana. Archaeocyantit levisivät tuolloin ympäri planeettaa. Vaiheen eliöstöä tunnetaan Kanadan Kalliovuorilla sijaitsevasta Burgess Shalen fossiiliesiintymästä, josta on löydetty muun muassa tikaripyrstön kaltainen, merenpohjalla saalistanut petoeläin Marrella splendens[9].
Kambrikauden lajiräjähdys
[muokkaa | muokkaa wikitekstiä]- Pääartikkeli: Kambrikauden lajiräjähdys
Kambrikausi oli suuri harppaus eliöstön kannalta prekambrin jälkeen. Fossiiliaineistosta päätellen lajisto pysyi suunnilleen samana kambrikauden alkuvaiheessa noin 15–20 miljoonaa vuotta, kunnes selkäjänteiset ja monet selkärangattomat, muun muassa piikkinahkaiset, lonkerojalkaiset ja niveljalkaiset ilmestyivät[10]. Tämän sanotaan johtuneen eliöille kehittyneestä kovasta suojakuoresta sekä eliöiden koon kasvamisesta. Kambrikaudella kehittyivät ensimmäiset nykyaikaiset eliöyhteisöt, kun kasvin- ja planktonsyöjien rinnalle tulivat saaliit sekä pedot. Aikakauden nopean evoluution uskotaan johtuneen myös eliöiden yksinkertaisesta perimästä, jossa suurikaan mutaatio ei tuottanut lisääntymis- tai elinkyvytöntä jälkeläistä. Suojakuorten kehitys oli luultavasti seurausta kauden alkupuolella ilmestyneistä pedoista. Koska niitä ei ollut merkittävästi prekambrin aikana, evoluutiolla ei ollut ollut tarvetta kehittää suojakuoria. Kuorellinen eliö säilyi hengissä kuoretonta paremmin.
Niin kutsuttu kambrikauden lajiräjähdys tapahtui noin 550–530 miljoonaa vuotta sitten. Lajien määrä yli kymmenkertaistui 10–15 miljoonan vuoden aikana. Lajiräjähdyksen esihistoria on osin hämärän peitossa. Sen arvellaan ulottuvan 750–570 miljoonan vuoden päähän. Erään analyysin mukaan 20 pääjaksosta 11 ilmestyi kambrikaudella, kahdeksan kambrikauden jälkeen ja yksi ennen kambrikautta. Molekyylitutkimusten mukaan jopa kuusi pääjaksoa olisi kehittynyt ennen kambrikautta. Erään käsityksen mukaan monisoluisten eliöiden kehitys olisi alkanut kasvaa 650 miljoonaa, toisen käsityksen mukaan jo 1 000 miljoonaa vuotta sitten. Jotkut väittävät monisoluisten perusjaon alkusuisiin ja jälkisuisiin tapahtuneen jo 1 200 miljoonaa vuotta sitten. Kambrikauden lajiräjähdys alkoi joidenkin tutkijoiden mielestä ainakin 30 miljoonaa vuotta ennen kambrikauden alkamista. Useimmat sienieläimet ilmestyivät kambrikauden alussa tai juuri ennen sitä. Ainakin matoja sekä meduusoita esiintyi ennen kambrikautta.
Kambrikaudella alkoi paleotsooinen maailmankausi ja fanerotsooinen aioni. Kambrikauden lajiräjähdys ajoitetaan varhaiskambriin, joka oli noin 542 ± 0,3 – 513 ± 2 miljoonaa vuotta sitten. Kambrikauden ajoituksesta on kuitenkin epävarmuutta ainakin 20 miljoonaa vuotta. Kambrikauden lajiräjähdys on selitettävissä evoluution todellisella käynnistymisellä ja eliömäärän kylmän jääkauden jälkeisellä lisääntymisellä kyseisenä aikana sekä kovakuoristen lajien, kuten trilobiittien, runsailla fossiilijäämillä. Suuria fossiilimääriä saattavat selittää ilmastonmuutoksen luomat olot, joissa fossiloituminen kävi helpommin. Kambrikauden aloittaa eliöstöjaotteluissa Tommot-vaihe, joka kesti arviolta kolme miljoonaa vuotta. Niveljalkaiset ovat kehittyneet luultavasti valejalkaisista lobopodeista[11].
Molekyylikellon mukaan ensimmäiset maakasvit olisivat eläneet jo kambrikaudella noin 500 miljoonaa vuotta sitten, mutta tätä ei kyetä suoraan fossiilein todistamaan.[12]
Kambrikaudella ilmestyneitä pääjaksoja
[muokkaa | muokkaa wikitekstiä]Kambrikauden aikana väitetään ilmestyneen 37, jopa 100 pääjaksoa. Keskikambrilta tunnetaan kanadalaisen fossiiliesiintymän Burgess Shalen perusteella seuraavat pääjaksot:
- Annelida (nivelmadot)
- Arthopoda (niveljalkaiset)
- Chordata (selkäjänteiset)
- Coelenterata (onteloeläimet)
- Echinodermata (piikkinahkaiset)
- Mollusca (nilviäiset)
- Onychophora (käsnäjalkaiset)
- Porifera (sienieläimet)
- Priapulida (makkaramadot)
Kambrikauden jako
[muokkaa | muokkaa wikitekstiä]Artikkelin alussa on kansainvälisen kerrostumajärjestön hyväksymä nykyinen kambrikauden jako. Yksi aiemmista jaoista on ollut seuraava:
- Yläkambri: 501–488 miljoonaa vuotta sitten
- kauden nimeämätön päätösvaihe: 496?–488 miljoonaa vuotta sitten
- Paibi: 501–496? miljoonaa vuotta sitten
- Keskikambri: 513–501 miljoonaa vuotta sitten
- Mayan: 502–501 miljoonaa vuotta sitten
- Amgan: 513–502 miljoonaa vuotta sitten
- Alakambri: 542–513 miljoonaa vuotta sitten
- Toyonian: 519–513 miljoonaa vuotta sitten
- Botomian: 524–519 miljoonaa vuotta sitten
- Atdaba: 530–524 miljoonaa vuotta sitten
- Tommot: 534–530 miljoonaa vuotta sitten
- Nemakit-Daldyn: 542–534 miljoonaa vuotta sitten
Katso myös
[muokkaa | muokkaa wikitekstiä]Lähteet
[muokkaa | muokkaa wikitekstiä]- ↑ Gradstein et al 2004
- ↑ Cambrian Period (englanniksi) Encyclopædia Britannica Online. Viitattu 24.11.2022.
- ↑ a b kambri | kambrikausi Tieteen termipankki. Viitattu 24.11.2022.
- ↑ https://web.archive.org/20031002003517/hometown.aol.com/darwinpage/paleontology.htm
- ↑ http://media-2.web.britannica.com/eb-media/92/792-004.gif
- ↑ http://media-3.web.britannica.com/eb-media/94/794-004.gif
- ↑ http://www.scotese.com/images/514.jpg
- ↑ Odontogriphus Museumkennis. Viitattu 27. toukokuuta 2007. (hollanniksi)
- ↑ Stephen Jay Gould, Ihmeellinen elämä , Art House, sivu 227 ja sivu 223
- ↑ Sean B. Carroll, Loputtomat kauniit muodot, Terra Cognita,Hakapaino Helsinki 2007, ISBN 978-952-5697-03-2, Eläinten evoluution suuri räjähdys: Kambrin räjähdys: niin monia niveljalkaisia, niin vähän aikaa, sivu 149
- ↑ Loputtomat kauniit muodot sivu 149
- ↑ Thomas Servais, Borja Cascales-Miñana, Christopher J. Cleal, Philippe Gerrienne, David A. T. Harper, Mareike Neumann: Revisiting the Great Ordovician Diversification of land plants: Recent data and perspectives. Palaeogeography, Palaeoclimatology, Palaeoecology, 15.11.2019, nro 534, s. 109280. doi:10.1016/j.palaeo.2019.109280 ISSN 0031-0182 Artikkelin verkkoversio. (englanniksi)
{kind=link}
{kind=link}
{kind=link}
Aiheesta muualla
[muokkaa | muokkaa wikitekstiä] Kuvia tai muita tiedostoja aiheesta Kambrikausi Wikimedia Commonsissa
Kuvia tai muita tiedostoja aiheesta Kambrikausi Wikimedia Commonsissa
Prekambri (4 567–541 miljoonaa vuotta sitten) | |||||||
|---|---|---|---|---|---|---|---|
| Hadeeinen aioni (4 567–4 000 mvs) | |||||||
| Arkeeinen aioni (4 000–2 500 mvs) | eoarkeeinen (4 000–3 600 mvs) • paleoarkeeinen (3 600–3 200 mvs) • mesoarkeeinen (3 200–2 800 mvs) • neoarkeeinen (2 800–2 500 mvs) | ||||||
| Proterotsooinen aioni (2 500–541 mvs) | paleoproterotsooinen (2 500–1 600 mvs) • mesoproterotsooinen (1 600–1 000 mvs) • neoproterotsooinen (1 000–541 mvs) | ||||||
541 miljoonaa vuotta sitten – nykyaika | |||||||
| Fanerotsooinen aioni (541 mvs – nykyaika) |
| ||||||